
153-2012-0258
在哺乳动物中,提出了一种已得到广泛认可的“二次打击”假说来证明 NASH 的发病机制(Day 和 James引用1998 年)。肝脏中的脂质积累或单纯脂肪变性被称为“第一次打击”,随后是“第二次打击”,这主要是由氧化应激和细胞因子释放增加引起的。肝脏脂肪变性不仅被认为是首发,而且被视为各种不同有害因素的诱发因素(Cortez-Pinto 等人,2008 年)。引用2006 年)。因此,“两次命中”假设已变为“多次平行命中”假设(Tilg 和 Moschen引用2010 年),其中胰岛素抵抗 (IR) 被视为第一步,这反过来又会导致高胰岛素血症和肝脂肪变性。最后,由于氧化和炎症应激以及脂质过氧化增强而发生肝损伤(Day引用2002 年)。
根据进化树,水禽和哺乳动物之间的遗传距离很大,并且它们的脂质代谢存在一些差异。例如,哺乳动物脂质合成的主要部位是脂肪组织,而鹅、鸭的脂质合成则集中在肝脏。哺乳动物的脂肪组织是脂质储存的主要场所,而肝脏和脂肪组织都是水禽的重要储存组织。哺乳动物的脂质储存大多是饱和的,而鹅或鸭的脂质主要在肝脏中是不饱和的。哺乳动物中的大部分脂质是外源性的,来源于食物,而水禽中的脂质大部分是内源性的,在肝脏中合成。此外,与哺乳动物相比,鸟类的血糖水平往往更高,这可能使它们比哺乳动物更能耐受高血糖。因此,哺乳动物脂肪肝的形成机制不同于水禽。
尽管对水禽脂肪肝形成机制的研究还处于初步阶段,但已经取得了一些进展(图1)。
图1.水禽脂肪肝形成过程及肝脂肪变性的保护机制。大写字母符号:I,胰岛素;IR,胰岛素受体;FA,脂肪酸;FFA,游离脂肪酸;ERS,内质网应激;TG,甘油三酯;VLDL,极低密度脂蛋白;HDL,高密度脂蛋白;HDL-R,HDL受体;IDL,中密度脂蛋白;IDL-R,IDL受体;HMG-COA,β-羟基-β-甲基戊二酰辅酶A;TCA,三羧酸循环。虚线表示:尽管 ERS 有助于 IR,但由过度喂养引起的鹅或鸭的 IR 和肝脂肪变性与 ERS 无关。红色和绿色斜体字母分别代表肝脏基因表达水平的降低和升高,当比较过量喂食富含碳水化合物的饮食的鹅或鸭与喂食正常饮食的对照组时。fatp,脂肪酸转运蛋白;pksG,羟甲基戊二酰-COA合酶;lpl,脂蛋白脂肪酶;scd,硬脂酰辅酶 A 去饱和酶;acly,ATP 柠檬酸裂合酶;dgat2,二酰基甘油 O-酰基转移酶 2;cs, 柠檬酸合酶; Apo-B,载脂蛋白
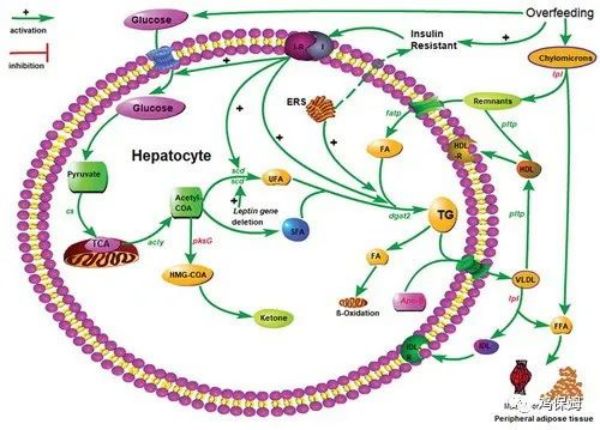
简而言之,在强制喂食后,鹅或鸭接受高能量饲料,其结果是肝脏中用于脂肪酸合成的底物(葡萄糖)大幅增加。同时,产生的甘油三酯含量远远超过载脂蛋白的转运能力,脂肪酸超过被β-氧化降解的脂肪酸,从而导致脂肪在肝脏中堆积。这增加了肝细胞的数量并促进了增殖(Herault 等人。引用2019 年),这会导致脂肪肝的形成,其重量可能是正常肝脏的 10 倍(Fournier 等人。引用1997 年)。脂质直接在肝脏中合成和沉积的方式是鹅和鸭脂肪肝形成的主要原因。此外,肝脏中产生的脂质主要由极低密度脂蛋白 (VLDL) 转运,而饮食中产生的脂质主要由乳糜微粒转运 (Griffin et al.引用1987 年)。由 VLDL 运输的大部分乳糜微粒和脂质在被血液中存在的脂蛋白脂肪酶 (lpl) 水解后沉积在外周脂肪组织和肌肉中。然而,如果这些脂蛋白转运的脂质不被lpl水解,就会在肝脏中特殊的脂蛋白受体的介导下被转运到肝脏沉积,从而促进脂肪肝的形成(Mulder et al.引用1993 年)。当肝脂肪变性发生时,过量喂养的鹅或鸭血清中的胰岛素水平维持在较高水平(Han et al.引用2008年;皮奥切等。引用2020 年)。体外实验表明,鹅肝细胞在接受葡萄糖或胰岛素处理后,可以快速生长并在短时间内繁殖(Wei 等人,2007 年)。引用2017 年)。因此,一般认为,鹅或鸭脂肪肝的形成,主要是由于肝脏脂质合成、转运和脂肪酸β-氧化失衡. 该过程与内质网应激 (ERS)、IR 和肝细胞生长和增殖密切相关。
胰岛素在碳水化合物和脂质代谢的调节中起着至关重要的作用。肝脏脂肪酸氧化、脂肪生成和蛋白质合成受胰岛素调节(O'Brien 和 Granner引用1996 年)。IR 意味着,当外周组织胰岛素变得不那么敏感时,身体需要更多的胰岛素来维持正常的血糖水平。内质网(ER)是真核细胞中负责蛋白质合成、折叠、组装和转运的重要细胞器。在哺乳动物中,IR 在非酒精性脂肪性肝病的发展中发挥作用,而 ERS 有助于 IR。胰岛素可能对脂质积累、脂肪生成基因的表达、肝脏中脂肪酸的氧化产生显着影响(Gontier 等人,2017 年)。引用2013),鹅原代肝细胞中VLDL-TG的组装和分泌(Han et al.引用2015 年)。正如 Geng 等人所证明的那样。(引用2015 ), 过度喂养导致 IR 和 ERS 有助于鹅的 IR, 表明 ERS 和 IR 参与鹅脂肪肝发展的机制。与哺乳动物相比,当伴有脂肪肝时,葡萄糖耐受不良/IR 似乎较少被诱导,因为在葡萄糖耐量试验期间,对照和处理鹅之间的血浆葡萄糖浓度差异小于对照组的三分之一小鼠,尽管小鼠注射葡萄糖的剂量比鹅高 2 倍 (Geng et al.引用2015 年)。这表明 IR 并不是鹅脂肪肝发展的唯一机制,并且有一个潜在的系统可以保护鹅免受 IR 和其他相关代谢紊乱的影响。弄清楚这个系统可能会为人类 NAFLD 的治疗提供一种治疗方法。
在哺乳动物中,高能量食物的消耗会引起肝脏 ERS (Ren et al.引用2013 年)。在原代肝细胞中,与高葡萄糖和棕榈酸酯一起孵育会增加GRP78的表达,这是 ERS 标记基因,尽管这在体内尚未观察到。例如,将膳食糖作为补充剂不能诱导肝 ERS (Geng et al.引用2016 年)。同样,过度饲喂 2 至 3 周会导致鹅出现 IR 和脂肪肝,但不会诱发 ERS(Geng 等人,2016 年)。引用2015 年),表明导致鹅 IR 和肝脂肪变性的过度饲喂与 ERS 无关。有人提出可能有其他机制诱导鹅的 IR 和肝脂肪变性。因此,值得探讨保护鹅肝脏免受 ERS 影响的因素,以及 IR 的非 ERS 机制。如果能充分理解相关机制,可以在鹅肝生产中降低超饲强度,有利于动物福利。
高通量测序结果显示,参与肝细胞脂肪酸合成的主要酶的基因表达水平显着升高,包括乙酰辅酶 A 羧化酶 ( acc )、脂肪酸合酶 ( fasn ) 、极长-链脂肪酸蛋白 6 ( elovl6 )、硬脂酰辅酶 A 去饱和酶 ( scd )、脂肪酸去饱和酶 1 ( fads1 ) (Lu et al.引用2015 年)。然而,lpl 和羟甲基戊二醇辅酶 A 合酶 ( pksG )的活性作为参与肝脏胆固醇合成的第一个关键酶,以及载脂蛋白 ( Apo-B ) 和lpl的基因表达水平,参与脂质包装和释放的关键基因,在过量喂食的鹅的肝脏中显着减少。同时,负责将外源脂质转运到细胞中的脂肪酸转运蛋白( fatp ) 基因的mRNA表达显着增强(Liu 等人,2016 年)。引用2016 年)。此外,过度喂养会导致脂质转运蛋白基因脂肪酸结合蛋白 4 ( FABP4;Even 等人引用2018 年)。脂肪组织中较高的A-FABP表达水平和A-FABP mRNA丰度与鹅的肝脏重量、肝重比、TG含量、血浆中VLDL浓度和HDL浓度密切相关(Liu et al.引用2019 年)。上调和下调的基因协同促进鹅或鸭在超饲早期脂肪肝的形成。正如之前的研究所揭示的那样,lpl 在来自细胞外乳糜微粒或 VLDL 的脂肪酸的脂解中起着重要作用,然后可以使用或沉积在脂肪或肌肉组织中(Saez 等人。引用2009 年;许等。引用2010年;安娜贝尔等人。引用2017 年)。lpl活性的降低增加了细胞外脂质扩散到肝细胞中的趋势。这些结果表明,水禽脂肪肝形成的机制主要归因于细胞质中新合成的内源性脂质和外源性脂质的储存和分泌之间的不平衡。肝脏脂质分泌能力不太可能抵消新合成的细胞质脂质的储存,导致鹅或鸭肝脏中的脂质沉积。
已有报道称,microRNA可能在鹅脂肪肝的形成中发挥作用。研究人员表明,miR-122在鹅的肝脏中高表达,并且其表达因过度喂食而下调 (Zhang et al.引用2018 年)。一些与脂质代谢相关的基因,包括脯氨酰 4-羟化酶亚基 α1、醛缩酶、果糖二磷酸 B 和丙酮酸激酶被预测并验证为鹅 miR- 122的靶基因(Zhang et al.引用2018 年)。高通量测序结果显示aan-miR-222和aan-miR-203a上调,而aan-miR-30d、aan-miR-125b-5p和aan-miR-146a-5p下调-在鹅的脂肪肝中调节 (Chen et al.引用2017 年)。刘等人。(引用2016 ) 报道了预测目标mRNA的通路分析表明,肝脏转录组中富集的通路几乎与 miRNA 分析的通路重叠。这说明 miRNA 在塑造肝脏转录组景观中起着重要作用,由此将与鹅肝形成相关的 miRNA 与其靶基因结合形成网络,可以从新的角度研究鹅肝形成的机制。
过度喂养显着增加了鹅或鸭血清中葡萄糖、总胆固醇 (TC)、甘油三酯和游离脂肪酸的浓度 (Lu et al.引用2015 年;安娜贝尔等人。引用2017 年)。鹅或鸭肝中新合成的脂质可以在肝细胞中以细胞质液滴的形式储存,也可以以脂蛋白的形式分泌到血液中(Hermier 等人,2007 年)。引用1991 年;埃罗等人。引用2010 年)。根据之前的一份报告,过度喂养大大增加了鸭或鹅的脂质参数、胰岛素和葡萄糖的血浆浓度。同时,VLDL 和高密度脂蛋白 (HDL) 的分泌率显着增加。肝脏合成甘油三酯的速度高于极低密度脂蛋白的分泌和脂肪酸β-氧化,这导致肝脏中大量的甘油三酯沉积(Han et al.引用2008 年)。结合起来,这些结果表明,在过度喂养后,肝脏中 TG 合成的速率显着高于 VLDL 分泌速率(Tavernier 等人,2016 年)。引用2017 年;刘等人。引用2019 年)。而且体内TG的转运通道饱和,血液中TG供应充足,保证了肝脏脂肪组织的快速形成。
血液参数被认为是疾病发生与否的指标之一。在肝脏炎症评估方面,传统的肝酶,如天冬氨酸转氨酶(AST)、丙氨酸转氨酶(ALT),已被临床应用于评估肝细胞损伤,但它们排除了NASH(Kunde et al.引用2005 年)。血液 TG、TC、HDL、ALT 和 AST 的升高通常与哺乳动物的脂肪肝有关。同时,谷氨酸丙酮酸转氨酶(GPT)和谷氨酸草乙酸转氨酶(GOT)的活性显着增加(Locsmandi 等引用2007 年)。过度喂养的鹅血清酶的这些变化与非酒精性脂肪肝患者的血清酶变化相似(Hermier 等人,2016 年)。引用2003 年)。血液生化显示,超饲鹅的 ALT 和 AST 水平明显高于对照组(Liu et al.引用2016 年)。慢性肝炎时,GOT/GPT值小于1,GOT/GPT值大于1时,肝细胞损伤严重,GPT活性随着慢性肝炎的发展而逐渐升高。文等。(引用2012 ) 报道当 GOT/GPT 值小于 1 时,过饲鸭的 GPT 逐渐增加。然而,肝细胞结构保持完好,肝细胞既没有坏死也没有破裂,这表明尽管发生了慢性肝炎,但肝细胞避免了鹅或鸭的过度喂养对肝脏的严重损伤,支持了一种保护机制的存在在水鸟的肝脏中。
